Mechanical and Electrical Responses in Living Matter

Extracted from Response in the Living and the Non-living.
1. THE MECHANICAL RESPONSE OF LIVING SUBSTANCES
Mechanical response – Different kinds of stimuli – Myograph – Characteristics of the response curve: period, amplitude, form – Modification of response curves.
One of the most striking effects of external disturbance on certain types of living substance is a visible change of form. Thus, a piece of muscle when pinched contracts. The external disturbance which produced this change is called the stimulus. The body which is thus capable of responding is said to be irritable or excitable. A stimulus thus produces a state of excitability which may sometimes be expressed by change of form.
Mechanical response to different kinds of stimuli
This reaction under stimulus is seen even in the lowest organisms; in some of the amœboid rhizopods, for instance. These lumpy protoplasmic bodies, usually elongated while creeping, if mechanically jarred, contract into a spherical form.
If, instead of mechanical disturbance, we apply salt solution, they again contract, in the same way as before. Similar effects are produced by sudden illumination, or by rise of temperature, or by electric shock.
A living substance may thus be put into an excitatory state by either mechanical, chemical, thermal, electrical, or light stimulus. Not only does the point stimulated show the effect of stimulus, but that effect may sometimes be conducted even to a considerable distance.
This power of conducting stimulus, though common to all living substances, is present in very different degrees. While in some forms of animal tissue irritation spreads, at a very slow rate, only to points in close neighbourhood, in other forms, as for example in nerves, conduction is very rapid and reaches far.
The visible mode of response by change of form may perhaps be best studied in a piece of muscle. When this is pinched, or an electrical shock is sent through it, it becomes shorter and broader. A responsive twitch is thus produced. The excitatory state then disappears, and the muscle is seen to relax into its normal form.
Mechanical lever recorder
In the case of contraction of muscle, the effect is very quick, the twitch takes place in too short a time for detailed observation by ordinary means.
A myographic apparatus is therefore used, by means of which the changes in the muscle are self-recorded. Thus we obtain a history of its change and recovery from the change.
The muscle is connected to one end of a writing lever. When the muscle contracts, the tracing point is pulled up in one direction, say to the right. The extent of this pull depends on the amount of contraction. A band of paper or a revolving drum-surface moves at a uniform speed at right angles to the direction of motion of the writing lever. When the muscle recovers from the stimulus, it relaxes into its original form, and the writing point traces the recovery as it moves now to the left, regaining its first position. A curve is thus described, the rising portion of which is due to contraction, and the falling portion to relaxation or recovery. The ordinate of the curve represents the intensity of response, and the abscissa the time (Figure 1).
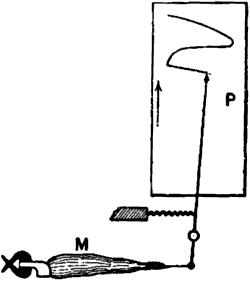
Fig. 1 – Mechanical lever recorder
The muscle M with the attached bone is securely held at one end, the other end being connected with the writing lever. Under the action of stimulus the contracting muscle pulls the lever and moves the tracing point to the right over the travelling recording surface P. When the muscle recovers from contraction, the tracing point returns to its original position. See on P the record of muscle curve.
Characteristics of the response curve: period, amplitude, and form
Just as a wave of sound is characterised by its (1) period, (2) amplitude, and (3) form, so may these response curves be distinguished from each other. As regards the period, there is an enormous variation, corresponding to the functional activity of the muscle. For instance, in tortoise it may be as high as a second, whereas in the wing muscles of many insects it is as small as 1/300 part of a second.
“It is probable that a continuous graduated scale might, as suggested by Hermann, be drawn up in the animal kingdom, from the excessively rapid contraction of insects to those of tortoises and hibernating dormice.” (1)
Differences in form and amplitude of curve are well illustrated by various muscles of the tortoise. The curve for the muscle of the neck, used for rapid withdrawal of the head on approach of danger, is quite different from that of the pectoral muscle of the same animal, used for its sluggish movements.
Again, progressive changes in the same muscle are well seen in the modifications of form which consecutive musclecurves gradually undergo. In a dying muscle, for example, the amplitude of succeeding curves is continuously diminished, and the curves themselves are elongated. Numerous illustrations will be seen later, of the effect, in changing the form of the curve, of the increased excitation or depression produced by various agencies.
Thus these response records give us a means of studying the effect of stimulus, and the modification of response, under varying external conditions, advantage being taken of the mechanical contraction produced in the tissue by the stimulus. But there are other kinds of tissue where the excitation produced by stimulus is not exhibited in a visible form. In order to study these we have to use an altogether independent method; the method of electric response.
Footnotes
1. Biedermann, Electro-physiology, p. 59.
2. ELECTRIC RESPONSE
Conditions for obtaining electric response – Method of injury – Current of injury – Injured end, cuproid: uninjured, zincoid – Current of response in nerve from more excited to less excited – Difficulties of present nomenclature – Electric recorder – Two types of response, positive and negative – Universal applicability of electric mode of response – Electric response a measure of physiological activity – Electric response in plants.
Unlike muscle, a length of nerve, when mechanically or electrically excited, does not undergo any visible change. That it is thrown into an excitatory state, and that it conducts the excitatory disturbance, is shown however by the contraction produced in an attached piece of muscle, which serves as an indicator.
But the excitatory effect produced in the nerve by stimulus can also be detected by an electrical method. If an isolated piece of nerve be taken and two contacts be made on its surface by means of non-polarisable electrodes at A and B, connection being made with a galvanometer, no current will be observed, as both A and B are in the same physico-chemical condition. The two points, that is to say, are iso-electric.
If now the nerve be excited by stimulus, similar disturbances will be evoked at both A and B. If, further, these disturbances, reaching A and B almost simultaneously, cause any electrical change, then, similar changes taking place at both points, and there being thus no relative difference between the two, the galvanometer will still indicate no current. This null-effect is due to the balancing action of B as against A. (See Figure 2a)
Conditions for obtaining electric response
If then we wish to detect the response by means of the galvanometer, one means of doing so will lie in the abolition of this balance, which may be accomplished by making one of the two points, say B, more or less permanently irresponsive. In that case, stimulus will cause greater electrical disturbance at the more responsive point, say A, and this will be shown by the galvanometer as a current of response. To make B less responsive we may injure it by means of a cross-sectional cut, a burn, or the action of strong chemical reagents.

Fig. 2 – Electric method of detecting nerve response
(a) Iso-electric contacts; no current in the galvanometer.
(b) The end B injured; current of injury from B to A: stimulation gives rise to an action current from A to B.
(c) Non-polarisable electrode.
Current of injury
We shall revert to the subject of electric response; meanwhile it is necessary to say a few words regarding the electric disturbance caused by the injury itself. Since the physicochemical conditions of the uninjured A and the injured B are now no longer the same, it follows that their electric conditions have also become different. They are no longer iso-electric. There is thus a more or less permanent or resting difference of electric potential between them. A current – the current of injury – is found to flow in the nerve, from the injured to the uninjured, and in the galvanometer, through the electrolytic contacts from the uninjured to the injured. As long as there is no further disturbance this current of injury remains approximately constant, and is therefore sometimes known as ‘the current of rest’ (Figure 2b).
A piece of living tissue, unequally injured at the two ends, is thus seen to act like a voltaic element, comparable to a copper and zinc couple. As some confusion has arisen, on the question of whether the injured end is like the zinc or copper in such a combination, it will perhaps be well to enter upon this subject in detail.
If we take two rods, of zinc and copper respectively, in metallic contact, and further, if the points A and B are connected by a strip of cloth s moistened with salt solution, it will be seen that we have a complete voltaic element. A current will now flow from B to A in the metal (Figure 3a) and from A to B through the electrolyte s. Or instead of connecting A and B by a single strip of cloth s, we may connect them by two strips s s′, leading to non-polarisable electrodes E E′. The current will then be found just the same as before, i.e. from B to A in the metallic part, and from A through s s′ to B, the wire W being interposed, as it were, in the electrolytic part of the circuit. If now a galvanometer be interposed at O, the current will flow from B to A through the galvanometer, i.e. from right to left. But if we interpose the galvanometer in the electrolytic part of the circuit, that is to say, at W, the same current will appear to flow in the opposite direction. In Figure 3c, the galvanometer is so interposed, and in this case it is to be noticed that when the current in the galvanometer flows from left to right, the metal connected to the left is zinc.
Compare Figure 3d, where A B is a piece of nerve of which the B end is injured. The current in the galvanometer through the non-polarisable electrode is from left to right. The uninjured end is therefore comparable to the zinc in a voltaic cell (i.e it is zincoid), the injured being copper-like or cuproid.(2)
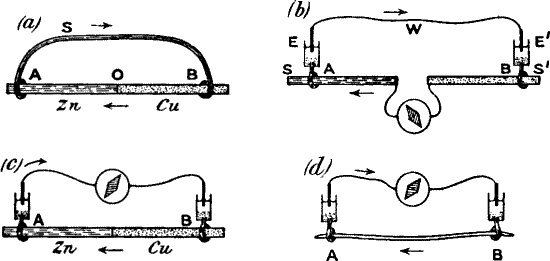
Fig. 3 – Diagram showing the correspondence between injured (b) and uninjured (a) contacts in nerve, and Cu and Zn in a voltaic element
Comparison of (c) and (d) will show that the injured end of B in (d) corresponds with the Cu in (c).
If the electrical condition of, say, zinc in the voltaic couple Figure 3c) undergoes any change (and I shall show later that this can be caused by molecular disturbance), then the existing difference of potential between A and B will also undergo variation. If for example the electrical condition of A approaches that of B, the potential difference will undergo a diminution, and the current hitherto flowing in the circuit will, as a consequence, display a diminution, or negative variation.
We have seen that a current of injury – sometimes known as ‘current of rest’ – flows in a nerve from the injured to the uninjured, and that the injured B is then less excitable than the uninjured A. If now the nerve be excited, there being a greater effect produced at A, the existing difference of potential may thus be reduced, with a consequent diminution of the current of injury. During stimulation, therefore, a nerve exhibits a negative variation. We may express this in a different way by saying that a ‘current of action’ was produced in response to stimulus, and acted in an opposite direction to the current of injury (Figure 2b). The action current in the nerve is from the relatively more excited to the relatively less excited.
Difficulties of present nomenclature
We shall deal later with a method by which a responsive current of action is obtained without any antecedent current of injury. ‘Negative variation’ has then no meaning. Or, again, a current of injury may sometimes undergo a change of direction (see note 4, p.30). In view of these considerations it is necessary to have at our disposal other forms of expression by which the direction of the current of response can still be designated.
Keeping in touch with the old phraseology, we might then call a current ‘negative’ that which flows from the more excited to the less excited. Or, bearing in mind the fact that an uninjured contact acts as the zinc in a voltaic couple, we might call it ‘zincoid,’ and the injured contact ‘cuproid.’
Stimulation of the uninjured end, approximating it to the condition of the injured, might then be said to induce a ‘cuproid change’.
The electric change produced in a normal nerve by stimulation may therefore be expressed by saying that there has been a negative variation, or that there was a current of action from the more excited to the less excited, or that stimulation has produced a cuproid change.
The excitation, or molecular disturbance, produced by a stimulus has thus a concomitant electrical expression. As the excitatory state disappears with the return of the excitable tissue to its original condition, the current of action will gradually disappear.(3) The movement of the galvanometer needle during excitation of the tissue thus indicates a molecular upset by the stimulus; and the gradual creeping back of the galvanometer deflection exhibits a molecular recovery.
This transitory electrical variation constitutes the ‘response,’ and its intensity varies according to that of the stimulus.
We have thus a method of obtaining curves of response electrically. After all, it is not essentially very different from the mechanical method. In this case we use a magnetic lever (Figure 4a), the needle of the galvanometer, which is deflected by the electromagnetic pull of the current, generated under the action of stimulus, just as the mechanical lever was deflected by the mechanical pull of the muscle contracting under stimulus.
The accompanying diagram (Figure 4b) shows how, under the action of stimulus, the current of rest undergoes a transitory diminution, and how on the cessation of stimulus there is gradual recovery of the tissue, as exhibited in the return of the galvanometer needle to its original position.

Fig. 4 – Electric recorder
(a) M muscle; A uninjured, B injured ends. E E′ non-polarising electrodes connecting A and B with galvanometer G. Stimulus produces ‘negative variation’ of current of rest. Index connected with galvanometer needle records curve on travelling paper (in practice, moving galvanometer spot of light traces curve on photographic plate). Rising part of curve shows effect of stimulus; descending part, recovery.
(b) O is the zero position of the galvanometer; injury produces a deflection A B; stimulus diminishes this deflection to C; C D is the recovery.
Two types of response – positive and negative
It may here be added that though stimulus in general produces a diminution of current of rest, or a negative variation (e.g. muscles and nerves), yet, in certain cases, there is an increase, or positive variation. This is seen in the response of the retina to light.
Again, a tissue which normally gives a negative variation may undergo molecular changes, after which it gives a positive variation. Thus Dr. Waller finds that whereas fresh nerve always gives negative variation, stale nerve sometimes gives positive; and that retina, which when fresh gives positive, when stale, exhibits negative variation.
The following is a summary of the two types of response:
1) Negative variation – Action current from more excited to less excited – cuproid change in the excited – e.g. fresh muscle and nerve, stale retina.
2) Positive variation – Action current from less excited to more excited – zincoid change in the excited – e.g. stale nerve, fresh retina.(4)
From this it will be seen that it is the fact of the electrical response of living substances to stimulus that is of essential importance, the sign plus or minus being a minor consideration.
Universal applicability of the electrical mode of response
This mode of obtaining electrical response is applicable to all living tissues, and in cases like that of muscle, where mechanical response is also available, it is found that the electrical and mechanical records are practically identical.
The two response curves seen in the accompanying diagram (Figure 5), and taken from the same muscle by the two methods simultaneously, clearly exhibit this. Thus we see that electrical response can not only take the place of the mechanical record, but has the further advantage of being applicable in cases where the latter cannot be used.
Electrical response: A measure of physiological activity
These electrical changes are regarded as physiological, or characteristic of living tissue, for any conditions which enhance physiological activity also, pari passu, increase their intensity. Again, when the tissue is killed by poison, electrical response disappears, the tissue passing into an irresponsive condition. Anæsthetics, like chloroform, gradually diminish, and finally altogether abolish, electrical response.
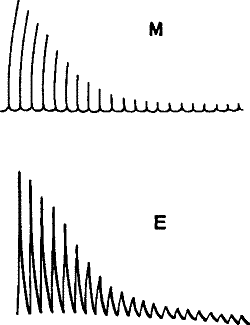
Fig. 5 – Simultaneous record of the mechanical (M) and (E) electrical responses of the muscle of frog (Waller)
From these observed facts – that living tissue gives response while a tissue that has been killed does not – it is concluded that the phenomenon of response is peculiar to living organisms.(5) The response phenomena that we have been studying are therefore considered as due to some unknown, super-physical ‘vital’ force and are thus relegated to a region beyond physical inquiry.
It may, however, be that this limitation is not justified, and surely, at least until we have explored the whole range of physical action, it cannot be asserted definitely that a particular class of phenomena is by its very nature outside that category.
Electric response in plants
But before we proceed to the inquiry as to whether these responses are or are not due to some physical property of matter, and are to be met with even in inorganic substances, it will perhaps be advisable to see whether they are not paralleled by phenomena in the transitional world of plants. We shall thus pass from a study of response in highly complex animal tissues to those given under simpler vital conditions.
Electric response has been found by Munck, Burdon-Sanderson, and others to occur in sensitive plants. But it would be interesting to know whether these responses were confined to plants which exhibit such remarkable mechanical movements, and whether they could not also be obtained from ordinary plants where visible movements are completely absent. In this connection, Kunkel observed electrical changes in association with the injury or flexion of stems of ordinary plants.(6)
My own attempt, however, was directed, not towards the obtaining of a mere qualitative response, but rather to the determination of whether throughout the whole range of response phenomena a parallel between animal and vegetable could be detected. That is to say, I desired to know, with regard to plants, what was the relation between intensity of stimulus and the corresponding response; what were the effects of superposition of stimuli; whether fatigue was present, and in what manner it influenced response; what were the effects of extremes of temperature on the response; and, lastly, if chemical reagents could exercise any influence in the modification of plant response, as stimulating, anæsthetic, and poisonous drugs have been found to do with nerve and muscle.
If it could be proved that the electric response served as a faithful index of the physiological activity of plants, it would then be possible successfully to attack many problems in plant physiology, the solution of which at present offers many experimental difficulties.
With animal tissues, experiments have to be carried on under many great and unavoidable difficulties. The isolated tissue, for example, is subject to unknown changes inseparable from the rapid approach of death. Plants, however, offer a great advantage in this respect, for they maintain their vitality unimpaired during a very great length of time.
In animal tissues, again, the vital conditions themselves are highly complex. Those essential factors which modify response can, therefore, be better determined under the simpler conditions which obtain in vegetable life.
In the following chapters it will be shown that the response phenomena are exhibited not only by plants but by inorganic substances as well, and that the responses are modified by various conditions in exactly the same manner as those of animal tissues. In order to show how striking are these similarities, I shall for comparison place side by side the responses of animal tissues and those I have obtained with plants and inorganic substances. For the electric response in animal tissues, I shall take the latest and most complete examples from the records made by Dr. Waller.
But before we can obtain satisfactory and conclusive results regarding plant response, many experimental difficulties will have to be surmounted. I shall now describe how this has been accomplished.(7)
Footnotes
2. In some physiological text-books much wrong inference has been made, based on the supposition that the injured end is zinc-like.
3. “The exciting cause is able to produce a particular molecular rearrangement in the nerve; this constitutes the state of excitation and is accompanied by local electrical changes as an ascertained physical concomitant.
The excitatory state evoked by stimulus manifests itself in nerve fibres by E.M. changes, and as far as our present knowledge goes by these only. The conception of such an excitable living tissue as nerve implies that of a molecular state which is in stable equilibrium. This equilibrium can be readily upset by an external agency, the stimulus, but the term “stable” expresses the fact that a change in any direction must be succeeded by one of opposite character, this being the return of the living structure to its previous state. Thus the electrical manifestation of the excitatory state is one whose duration depends upon the time during which the external agent is able to upset and retain in a new poise the living equilibrium, and if this is extremely brief, then the recoil of the tissue causes such manifestation to be itself of very short duration.” – Textbook of Physiology, ed. by Schäfer, ii. 453.
4. I shall here mention briefly one complication that might arise from regarding the current of injury as the current of reference, and designating the response current either positive or negative in relation to it. If this current of injury remained always invariable in direction – that is to say, from the injured to the uninjured – there would be no source of uncertainty. But it is often found, for example in the retina, that the current of injury undergoes a reversal, or is reversed from the beginning. That is to say, the direction is now from the uninjured to the injured, instead of the opposite. Confusion is thus very apt to arise. No such misunderstanding can however occur if we call the current of response towards the more excited positive, and towards the less excited negative.
Extracted from Response in the Living and the Non-living, published by A Distant Mirror.